
توالی یابی مکسام–گیلبرت روشی برای تعیین توالی دی ان ای است که الن مکسام و والتر گیلبرت در ۱۹۷۶–۱۹۷۷ ابداع کردند. این روش بر اساس تغییر شیمیایی جزئی خاص باز نوکلئوتیدی دی ان ای و پس از آن تسهیم پیوند ستون فقرات دی ان ای در نواحی های مجاور نوکلئوتیدهای تغییر یافته است. [ ۱]
مکسام–گیلبرت پراستفاده ترین روش برای توالی یابی پس از روش سنگر است که نماینده اولین نسل از روش های توالی یابی است. روش مکسام–گیلبرت دیگر به صورت گسترده استفاده نمی شود و روش توالی نسل بعدی جایگزین آن شده است.
گرچه مکسام و گیلبرت روش توالی یابی خود را دو سال پس از آن که فردریک سانگر و آلن کولسون توالی یابی مثبت - منفی[ ۲] [ ۳] را منتشر کردند، انتشار دادند اما توالی یابی مکسام–گیلبرت به سرعت محبوب تر شد چون دی ان ای خالص شده می توانست به صورت مستقیم استفاده شود در حالی که روش اولیه سنگر به کپی از شروع هر رید برای تولید تک رشته دی ان ای نیاز داشت. اما با بهبود روش پایانش زنجیره، استفاده از روش مکسام–گیلبرت به دلیل پیچیدگی های فنی که از استفاده از آن در کیت های استاندارد زیست شناسی مولکولی جلوگیری می کند، استفاده از مواد شیمیایی خطرناک و مشکلات توسعه آن کاهش یافت. [ ۴]
فرایند توالی یابی مکسام - گیلبرت به سه گام آماده سازی و پایانش شیمیایی و استنتاج رشته تقسیم می شود.
• مرحله آماده سازی: در این گام دو فعالیت عمده انجام می شود: یک - تک رشته ای کردن دی ان ای: از آن جایی که دی ان ای از دو رشته مکمل هم ساخته شده است، کافی است تا توالی یک رشته از آن مشخص شود تا رشته دوم از روی آن ساخته شود. این کار با گرم کردن دی ان ای و افزودن مجموعه ای از مواد شیمیایی از جمله گلیسرول انجام می گیرد. دو - افزودن گروه فسفات P32 که خاصیت رادیواکتیو دارد و در پرتونگاری ردپا از خود به جا می گذارد.
• مرحله پایانش شیمیایی: در این گام چهار نوع آزمایش انجام می شود که این آزمایش ها بخش های کوچکی از دی ان ای را تولید می کنند. به این ترتیب که رشته می تواند در باز مربوط به آن آزمایش شکسته شود. در آزمایش نوع اول مربوط به گروه های A+G، دوم مربوط به G سوم مربوط به C و چهارم مرتبط به C+Tاست. برای مثال افزودن نمک ( کلرید سدیم ) به واکنش هیدرازین سبب مهار واکنش تیمین برای واکنش C می شود، پورین ( A+G ) با استفاده از اسید فرمیک پورین زدایی می شود، گوانین ( و تا حدی آدنین ) توسط دی متیل سولفات متیله می شود و پیرامیدین ( C+T ) با استفاده از هیدرازین هیدرولیز می شود. در نتیجه مجموعه ای از برچسب قطعات تولید شده از پایان برچسب گذاری شده رادیواکتیوی تا اولین برش در هر مولکول تولید می شود.
• استنتاج رشته: قطعات در چهار واکنش در ژل آکریل آمید تفکیک و الکتروفورز می شوند تا به ترتیب اندازه جدا شوند. برای نمایش بخش ها، ژل در معرض اشعه Xقرار می گیرد تا اتورادیوگرافی انجام شود و نوارهای تیره ای را که بیانگر مکان مولکول رادیواکتیو شده است را نمایش دهد. از حضور یا غیبت برخی قسمت ها می توان رشته را استنباط کرد. [ ۱] [ ۵] به این ترتیب که اگر نواری در هر دو واکنش C+tو C باشد به این منظور است که در آن مکان باز C قرار دارد اگر تنها در C+t نواری داشته باشیم یعنی باز T خواهیم داشت. به صورت مشابه استدلال هایی برای بازهای Aو G وجود دارد.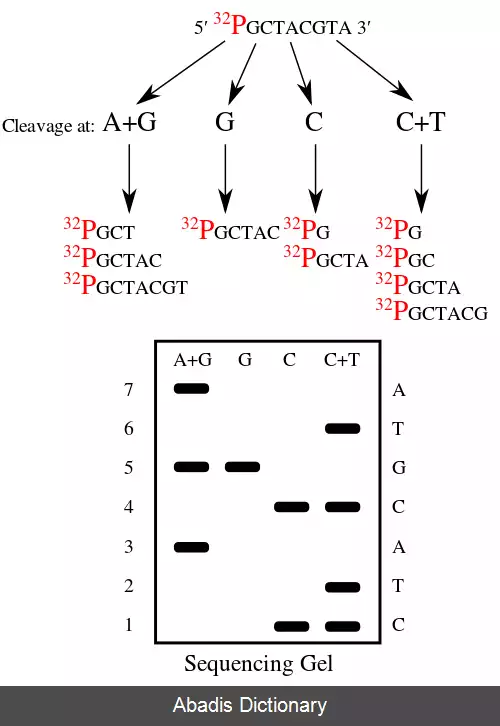
این نوشته برگرفته از سایت ویکی پدیا می باشد، اگر نادرست یا توهین آمیز است، لطفا گزارش دهید: گزارش تخلفمکسام–گیلبرت پراستفاده ترین روش برای توالی یابی پس از روش سنگر است که نماینده اولین نسل از روش های توالی یابی است. روش مکسام–گیلبرت دیگر به صورت گسترده استفاده نمی شود و روش توالی نسل بعدی جایگزین آن شده است.
گرچه مکسام و گیلبرت روش توالی یابی خود را دو سال پس از آن که فردریک سانگر و آلن کولسون توالی یابی مثبت - منفی[ ۲] [ ۳] را منتشر کردند، انتشار دادند اما توالی یابی مکسام–گیلبرت به سرعت محبوب تر شد چون دی ان ای خالص شده می توانست به صورت مستقیم استفاده شود در حالی که روش اولیه سنگر به کپی از شروع هر رید برای تولید تک رشته دی ان ای نیاز داشت. اما با بهبود روش پایانش زنجیره، استفاده از روش مکسام–گیلبرت به دلیل پیچیدگی های فنی که از استفاده از آن در کیت های استاندارد زیست شناسی مولکولی جلوگیری می کند، استفاده از مواد شیمیایی خطرناک و مشکلات توسعه آن کاهش یافت. [ ۴]
فرایند توالی یابی مکسام - گیلبرت به سه گام آماده سازی و پایانش شیمیایی و استنتاج رشته تقسیم می شود.
• مرحله آماده سازی: در این گام دو فعالیت عمده انجام می شود: یک - تک رشته ای کردن دی ان ای: از آن جایی که دی ان ای از دو رشته مکمل هم ساخته شده است، کافی است تا توالی یک رشته از آن مشخص شود تا رشته دوم از روی آن ساخته شود. این کار با گرم کردن دی ان ای و افزودن مجموعه ای از مواد شیمیایی از جمله گلیسرول انجام می گیرد. دو - افزودن گروه فسفات P32 که خاصیت رادیواکتیو دارد و در پرتونگاری ردپا از خود به جا می گذارد.
• مرحله پایانش شیمیایی: در این گام چهار نوع آزمایش انجام می شود که این آزمایش ها بخش های کوچکی از دی ان ای را تولید می کنند. به این ترتیب که رشته می تواند در باز مربوط به آن آزمایش شکسته شود. در آزمایش نوع اول مربوط به گروه های A+G، دوم مربوط به G سوم مربوط به C و چهارم مرتبط به C+Tاست. برای مثال افزودن نمک ( کلرید سدیم ) به واکنش هیدرازین سبب مهار واکنش تیمین برای واکنش C می شود، پورین ( A+G ) با استفاده از اسید فرمیک پورین زدایی می شود، گوانین ( و تا حدی آدنین ) توسط دی متیل سولفات متیله می شود و پیرامیدین ( C+T ) با استفاده از هیدرازین هیدرولیز می شود. در نتیجه مجموعه ای از برچسب قطعات تولید شده از پایان برچسب گذاری شده رادیواکتیوی تا اولین برش در هر مولکول تولید می شود.
• استنتاج رشته: قطعات در چهار واکنش در ژل آکریل آمید تفکیک و الکتروفورز می شوند تا به ترتیب اندازه جدا شوند. برای نمایش بخش ها، ژل در معرض اشعه Xقرار می گیرد تا اتورادیوگرافی انجام شود و نوارهای تیره ای را که بیانگر مکان مولکول رادیواکتیو شده است را نمایش دهد. از حضور یا غیبت برخی قسمت ها می توان رشته را استنباط کرد. [ ۱] [ ۵] به این ترتیب که اگر نواری در هر دو واکنش C+tو C باشد به این منظور است که در آن مکان باز C قرار دارد اگر تنها در C+t نواری داشته باشیم یعنی باز T خواهیم داشت. به صورت مشابه استدلال هایی برای بازهای Aو G وجود دارد.
