
کراسینگ اور[ ۱] [ ۲] [ ۳] یا چلیپایی شدن ( به انگلیسی: Crossing over ) می گویند. تبادل ماده ژنتیکی در طی تولید مثل جنسی بین کروماتیدهای غیر خواهر دو کروموزوم همولوگ است که منجر به کروموزوم های نوترکیب می شود. یکی از مراحل نهایی نوترکیبی ژنتیکی است که در مرحله پاکیتن پروفاز I میوز طی فرآیندی به نام سیناپسیس رخ می دهد. سیناپسیس قبل از ایجاد کمپلکس سیناپتونمال شروع می شود و تا پایان پروفاز I کامل می شود. کراسینگ اور در حقیقت آخرین مرحلهٔ نوترکیبی ژنی است. برای مجموعه ثابتی از شرایط ژنتیکی و محیطی، نوترکیب در یک ناحیه خاص از یک ساختار ( کروموزوم ) تمایل به ثابت بودن دارد و همین امر برای مقدار کراسینگ که در تولید نقشه های ژنتیکی استفاده می شود، صادق است. [ ۴] [ ۵]
در یوکاریوت ها از هر ژن دو نسخه حمل می شود که به آن الل ( دگره ) می گویند. هر یک از آن ها از یکی از پدر یا مادر به فرزند رسیده است. در مرحلهٔ گامت زایی و در بخش متافاز آن این الل ها از هم جدا شده و هر یک وارد یک گامت می شوند. در طی کراسینگ اور این الل ها می توانند تغییر کنند. [ ۶]
مورگان به طور دقیقی این فرایند را در مگس سرکه آزمایش کرد و به نتایج بزرگی رسید.
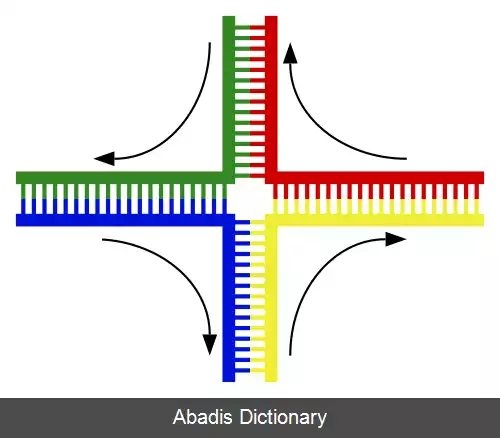
در آغاز یک شکست دورشته ای در دی ان ای توسط Spo۱۱ و در مرحله میوز I رخ می دهد. این پروتئین یک اگزونوکلئاز است؛ که با شکست در سر۵ٰ سبب ایجاد یک تک رشته با سر ۳ٰ می شود. در میوز عامل اصلی نوترکیبی Dmc۱ است. این پرونئین به همراه Rad۵۱ تک رشته ای ایجاد شده را می پوشانند و چیزی شبیه به فیلامنت ساخته می شود. در مرحلهٔ بعد هر رشته نقص خود را از روی کروماتید موجود اصلاح می کند و رشته کامل می شود. در این حالت چهارراهی حاصل می شود که به افتخار شخصی که آن را کشف کرده است آن را تقاطع هالیدی[ ۷] نامند که در حقیقت یک ساختمان چهارتایی از چهار رشته است.
نرخ نوترکیبی ژنی و در نهایت کراسینگ اوور در ژن های مختلف متفاوت است و بستگی به فاصلهٔ ژنها دراد. ژن های نزدیک به هم احتمال کراسینگ اور کمتری دارند. به وسیلهٔ نشانگر می توان این رویداد را بررسی کرد همچنین توالی دی ان ای را می توان توسط این نشانگرها بررسی کرده و در درمان بیماری های ژنتیکی استفاده کرد.
اگرچه کراسینگ اور یک روش مناسب برای نوترکیبی است اما می تواند توازن ژن ها را به هم بریزد و آن ها را از حالت متعادل خارج کند. به این شکل گامت های ایجاد شده دارای نقص هستند و قسمتی از ژنوم آن ها حذف یا افزوده شده است. دوبرابرشدگی ها و وارونه شدگی ها نیز نتیجهٔ چلیپایی شدن نابرابر است.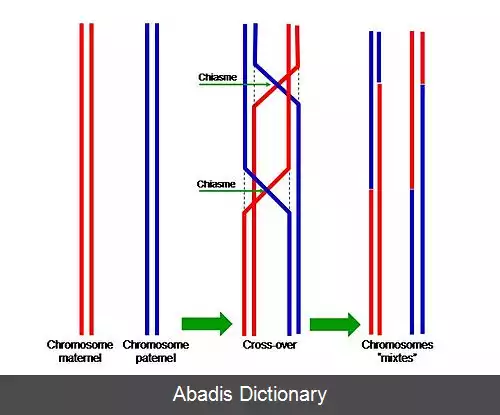
این نوشته برگرفته از سایت ویکی پدیا می باشد، اگر نادرست یا توهین آمیز است، لطفا گزارش دهید: گزارش تخلفدر یوکاریوت ها از هر ژن دو نسخه حمل می شود که به آن الل ( دگره ) می گویند. هر یک از آن ها از یکی از پدر یا مادر به فرزند رسیده است. در مرحلهٔ گامت زایی و در بخش متافاز آن این الل ها از هم جدا شده و هر یک وارد یک گامت می شوند. در طی کراسینگ اور این الل ها می توانند تغییر کنند. [ ۶]
مورگان به طور دقیقی این فرایند را در مگس سرکه آزمایش کرد و به نتایج بزرگی رسید.
در آغاز یک شکست دورشته ای در دی ان ای توسط Spo۱۱ و در مرحله میوز I رخ می دهد. این پروتئین یک اگزونوکلئاز است؛ که با شکست در سر۵ٰ سبب ایجاد یک تک رشته با سر ۳ٰ می شود. در میوز عامل اصلی نوترکیبی Dmc۱ است. این پرونئین به همراه Rad۵۱ تک رشته ای ایجاد شده را می پوشانند و چیزی شبیه به فیلامنت ساخته می شود. در مرحلهٔ بعد هر رشته نقص خود را از روی کروماتید موجود اصلاح می کند و رشته کامل می شود. در این حالت چهارراهی حاصل می شود که به افتخار شخصی که آن را کشف کرده است آن را تقاطع هالیدی[ ۷] نامند که در حقیقت یک ساختمان چهارتایی از چهار رشته است.
نرخ نوترکیبی ژنی و در نهایت کراسینگ اوور در ژن های مختلف متفاوت است و بستگی به فاصلهٔ ژنها دراد. ژن های نزدیک به هم احتمال کراسینگ اور کمتری دارند. به وسیلهٔ نشانگر می توان این رویداد را بررسی کرد همچنین توالی دی ان ای را می توان توسط این نشانگرها بررسی کرده و در درمان بیماری های ژنتیکی استفاده کرد.
اگرچه کراسینگ اور یک روش مناسب برای نوترکیبی است اما می تواند توازن ژن ها را به هم بریزد و آن ها را از حالت متعادل خارج کند. به این شکل گامت های ایجاد شده دارای نقص هستند و قسمتی از ژنوم آن ها حذف یا افزوده شده است. دوبرابرشدگی ها و وارونه شدگی ها نیز نتیجهٔ چلیپایی شدن نابرابر است.


wiki: کراسینگ اور